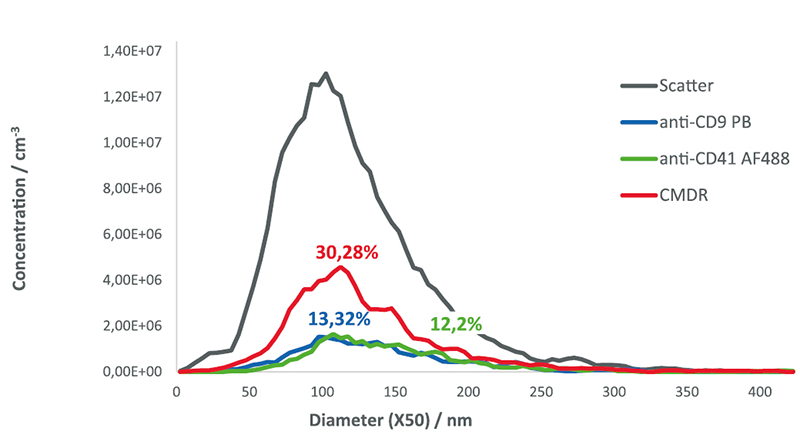
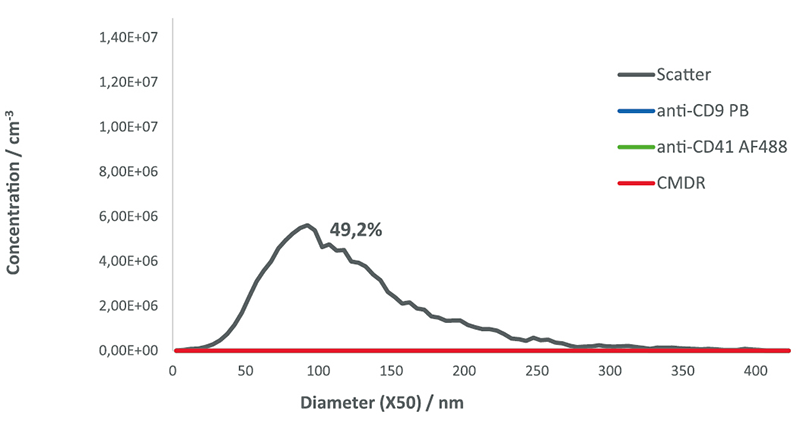
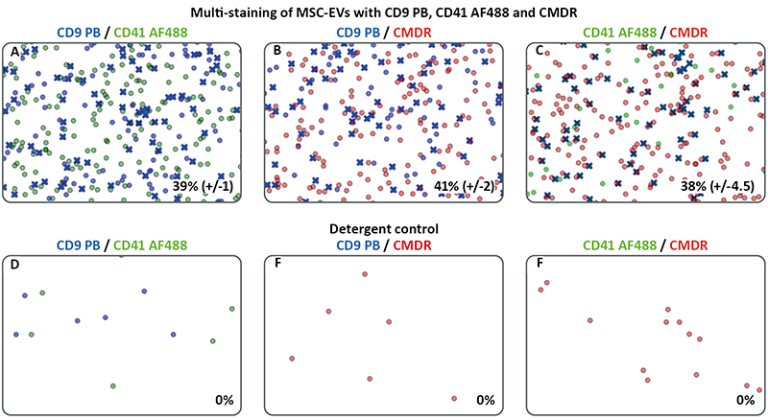
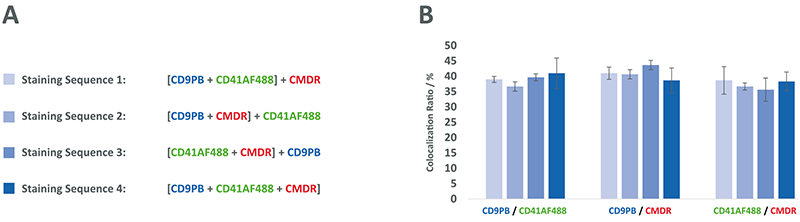
1. Koliha N, Heider U, Ozimkowski T, et al.:
Melanoma affects the composition of blood cell-derived extracellular vesicles [original research]. Front Immunol.
2016 Jul;7(282). English.
DOI:10.3389/fimmu.2016.00282
2. Koliha N, Wiencek Y, Heider U, et al.:
A novel multiplex bead-based platform highlights the diversity of extracellular vesicles. J Extracell Vesicles.
2016;016;5:29975. PubMed PMID: 26901056; PubMed
Central PMCID: PMCPMC4762227.
3. Wiklander OP, Nordin JZ, O’Loughlin A, et al.:
Extracellular vesicle in vivo bio-distribution is determined
by cell source, route of administration and targeting.
J Extracell Vesicles. 2015;4:26316. PubMed PMID:
25899407; PubMed
Central PMCID: PMCPMC4405624.
4. Wiklander OPB, Bostancioglu RB, Welsh JA, et al.:
Systematic methodological eva-luation of a multiplex
bead-based flow cytometry assay for detection of
extra-cellular vesicle surface signatures
Front Immunol. 2018 Jun;9(1326).
English. doi: 10.3389/fimmu.2018.01326.
5. Kowal J, Arras G, Colombo M, et al.:
Proteomic comparison defines novel markers to characterize
heterogeneous populations of extracellular vesicle
sub-types. Proc Natl Acad Sci U S A. 2016 Feb 8. PubMed
PMID: 26858453.
DOI:10.1073/pnas.1521230113.
6. Giebel B.
On the function and hetero-geneity of extracellular
vesicles. Ann Transl Med. 2017 Mar;5(6):150.
PubMed PMID: 28462230; PubMed Central PMCID:
PMCPMC5395490.
7. Witwer KW, Buzas EI, Bemis LT, et al.:
Standardization of sample collection, isolation and
analysis methods in extracellular vesicle research. J Extracell
Vesicles. 2013;2. PubMed PMID: 24009894; PubMed
Central PMCID: PMCPMC3760646.
DOI:10.3402/jev.v2i0.20360
8. Sharma S, Rasool HI, Palanisamy V, et al.:
Structuralmechanical characterization of nanoparticle
exosomes in human saliva, using correlative AFM, FESEM,
and force spectroscopy. ACS Nano. 2010;4(4):1921–1926